Biomolecular simulations to investigate neurotrophin receptor mechanisms and interactions with neurotrophin mimetics
|
Neurotrophins (NTs) are proteins with critical roles in neuronal survival, axonal and dendritic network maintenance, and synaptic plasticity (Gómez et al. 2013; Huang et al. 2001; Majdan et al. 2001). Because of their important functions, NTs and their receptors are pharmaceutical targets for neurodegenerative diseases (Meldolesi et al. 2017). In humans there are four NTs; nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3), neurotrophin-4/5 (NT-4/5), which bind to four NT receptors; tropomyosin receptor kinases A, B, C (TrkA, TrkB, TrkC), which are receptor tyrosine kinases, and p75 neurotrophin receptor (p75NTR), which is a tumor necrosis factor receptor (Chao et al. 2003). The NT receptors are transmembrane receptors that exert their activity intracellularly upon NT binding. Understanding the mechanism of receptor activation can aid the development of successful pharmaceuticals. Biomolecular simulations have been proven in the past to provide valuable insights into the structural and dynamical aspects of several transmembrane receptors by calculating the evolution of biomolecular systems with time (Arkhipov et al. 2017; Lelimousin et al. 2016; Prakaash et al. 2021). Since the compounds designed in the EuroNeurotrophin consortium act on NT receptors, simulations are employed to assist drug design. This article provides a short review of molecular simulations performed on NT receptors from the extracellular to the intracellular domains (Figure 2), as well as work done by the consortium members at Heidelberg Institute for Theoretical Studies (HITS).
The extracellular domain (Figure 1) of the NT receptors has been shown to be the binding site of NTs, (Gong et al. 2008; Wehram et al. 2007) as well as small-molecule receptor agonists such amitriptyline and BNN27 (Shoemark et al. 2015; Pediaditakis et al. 2016; Pediaditakis et al. Neuropharmacology, 2016). Saturation transfer difference nuclear magnetic resonance (STD-NMR) experiments have indicated the interface between NGF and the extracellular regions of TrkA as the binding region of BNN27 TrkA agonist, while docking and molecular dynamics (MD) simulations showed that the compound can be stabilized at the interface of the two proteins (Pediaditakis et al, Neuropharmacology, 2016). Additionally, MD simulations have shown that BNN27 binds favorably at the interface of p75NTR and NGF (Pediaditakis et al, Front. Pharmacol. 2016). Accordingly, BNN27 analogues developed in the EuroNeurotrophin consortium are examined for their ability to bind stably to the interface of NT and receptors, using docking and MD simulations. 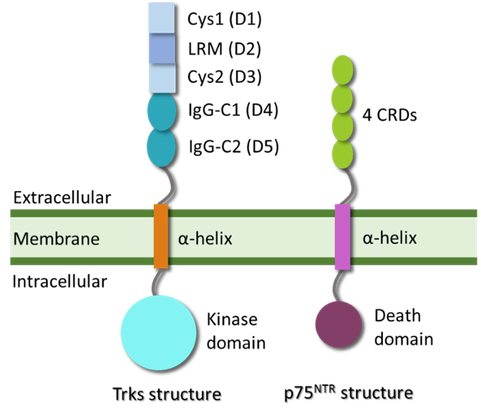
Figure 2. Structures of Trk and p75NTR receptors. The extracellular part of each Trk has 5 domains; two cysteine-rich domains (Cys1:D1, Cys2:D3), one leucine-rich motif (LRM:D2) and two immuno-globulin-like domains (Ig-C1:D4, Ig-C2:D5), while p75NTR has four cysteine-rich domains (CRDs) in the extracellular segment. Both types of receptors have a helical transmembrane domain and, intracellularly, a kinase or a death domain in the Trk and p75NTR, respectively. The structures of all the domains have been solved by X-ray crystallography or NMR except for the linkers between the transmembrane domain and the other domains.
The complex of the extracellular domain of TrkA dimer bound to NGF has been investigated through MD simulations for its stability and possible conformations. Previous MD simulations have indicated that the complex forms water-mediated hydrogen-bond interactions at its interface, (Settanni et al. 2003) while another study highlighted the importance of the N-terminal peptide of NGF for the binding to TrkA (Berrera et al. 2006). Advanced simulations have also been used to compute the free energy of different conformations of the full extracellular TrkA domain in complex with NGF (Pietropaolo et al. 2018). An important feature of the extracellular domain of the Trk receptors and p75NTR is its glycosylation state, which is necessary for membrane localization and ligand-dependent activation (Watson et al. 1999). In this regard, simulations that examine the conformational effects of glycosylation in the extracellular segments of the receptors have been employed within the consortium (Figure 3).
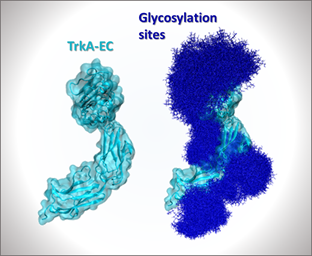
Figure 3: Extracellular (EC) domain of TrkA from crystral struc-ture PDB ID: 2IFG (Wehram et al. 2007). The protein is shown with (right) and without (left) glycans at the glycosylation sites. Various glycan conformations observed in MD simulation are superimposed and shown in dark blue with the protein surface in cyan. (Athanasiou et al, unpublished data).
NT binding to the extracellular domain leads to a conformational change to the intracellular domain that is mediated by the transmembrane domain. Understanding the dynamics of this domain is pivotal for elucidating the mechanism of receptor activation. Additionally, the transmembrane domain of TrkB has been shown to be a target of antidepressant drugs (Casarotto et al. 2021). MD simulations revealed that drug binding to the transmembrane domain of TrkB can stabilize the helical dimer in an arrangement that is independent of cholesterol concentration in the membrane (Casarotto et al. 2021). The interaction between the transmembrane domains of TrkA and p75NTR has also been studied recently, with simulations indicating that p75NTR interacts with TrkA in a way that leaves the active interface of TrkA accessible for another TrkA monomer, thus facilitating TrkA activation (Franco et al. 2021). Based on the aforementioned data, multiscale simulations are performed within the EuroNeurotrophin consortium to understand the dynamical behavior of the transmembrane homodimers of NT receptors (Figure 4), as well as their interactions with small molecules.

Figure 4: Different arrangements of the transmembrane (TM) helical TrkA homodimer obtained by coarse-grained simulations. The helical dimers have been aligned to the first helix shown in white. The second helix from the available NMR structure (Fran-co et al. 2020) is shown in orange and the one from different simulation snapshots in purple. (Athanasiou et al, unpublished data).
In the intracellular region of the receptors there are the juxtamembrane linker and globular domains. The latter is a kinase for the Trks and a death domain for the p75NTR. The juxtamembrane domains of the epidermal growth factor receptor (EGFR) and the fibroblast growth factor receptor 3 (FGFR3) play an important role in receptor activation (Tamagaki et al. 2014; Red Brewer et al. 2009). The juxtamembrane domain of TrkA has been probed with MD simulations for its interaction with the membrane (Wang et al. 2019). The simulations showed interactions of the basic protein residues with anionic membrane lipids and a three-residue insertion into the membrane (Wang et al. 2019). In order to investigate the relative orientation and interactions with lipids found in neuronal membranes, simulations of the transmembrane and intracellular domains of NT receptors (Figure 5) are being performed in the EuroNeurotrophin consortium, which have revealed specific interaction patterns, potentially important for receptor function.
Each of the aforementioned simulations provides a piece of the puzzle of understanding how these receptors function. Together with simulations of the complete receptors, they will offer valuable insight into the recognition and activation processes, which will aid the design of neuroactive compounds that will target these receptors to treat neurodegeneration. 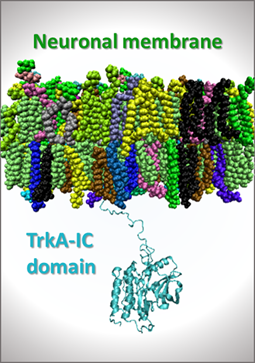
Figure 5: System of the transmembrane TrkA domain embedded in a neuronal membrane and the intracellular (IC) domain (cy-an) in the aqueous solvent region. (Athanasiou et al, un-published data).
|

Christina joined the EuroNeurotrophin network as a PhD student at the Heidelberg Institute for Theoretical Studies (HITS). She investigates the mechanism of action of neurotrophin small molecule mimetics and potentiators, through the use of molecular simulations and mathematical modelling techniques. She also studies the interactions between neurotrophin mimetics and their receptors, as well as conformational changes and allosteric effects caused by the small molecules.
|
